- Have any questions? Contact us!
- info@dr-rath-foundation.org


Plasmin-Induced Proteolysis and the Role of Apoprotein(a), Lysine, and Synthetic Lysine Analogs
October 4, 2017
Lipoprotein(a) Reduction by Ascorbate
October 4, 2017Solution to the Puzzle of Human Evolution

Matthias Rath M.D. (1992)
Journal of Orthomolecular Medicine 7: 73-80.
“Much light will be thrown on the origin of man and his history.”
Introduction
Until now human evolution has remained one of the greatest puzzles of mankind. Neither paleoanthropology nor behavioral or genetic approaches are able to explain the dramatic development that let to the evolution of modern man and made him the dominant species on earth. This explanation is provided by a fascinating combination of genetic, metabolic, environmental, and dietary elements. Several of my recent discoveries turned out to be important to solve the puzzle of human evolution.
After the loss of endogenous ascorbate synthesis in the ancestor of man, scurvy became the greatest threat to the evolutionary survival of our ancestors.¹ Apo(a) and Lp(a) became important metabolic constituents in man and subhuman primates after they had lost the ability for endogenous ascorbate synthesis.² Apo(a) functions as an adhesive protein and was a metabolic key in the development of intelligence and fertility during human evolution.³ These discoveries, together with the fact that the evolution of man was greatly accelerated during the Ice Ages starting about 2.5 million years ago,4 led me to the solution of the puzzle of human evolution, which will be presented in the following paragraphs.
The Loss of Endogenous Ascorbate Production – The Genetic Precondition For Human Evolution
About 40 million years ago the ancestor of man lost the ability to synthesize ascorbate endogenously. This was the result of the mutation of the gene encoding for the enzyme L-gulono-?-lactone oxidase, a key enzyme in the conversion of glucose to ascorbate. This genetic mutation left all descendants, including all human beings living today, dependent on sufficient exogenous ascorbate supply in the diet.
The precondition for this genetic mutation was a sufficient dietary supply of ascorbate. The precondition was met by the fact that at the time of the mutation our ancestors lived in the central regions of Africa and their diet consisted mainly of fruits and other nutrition rich in ascorbate and other vitamins. Nevertheless, as a result of this mutation the availability of ascorbate in the body of our ancestors dropped from between 10,000 to 20,000 milligrams synthesized endogenously every day, to several hundred milligrams taken up in the diet of the African habitat. More than 30 million years later this genetic defect was completely unmasked by environmental conditions triggering the evolution of man.
Ice Ages – The Environmental Trigger For the Evolution of Man
The evolution of man was greatly accelerated during the Ice Ages, which started about 2.5 million years ago. During this relatively short time the size of the human brain quadrupled and man became the dominant species on earth. Since that time the glaciation periods occurred periodically about every 100,000 years, lasting for several tens of thousands of years. During the short warm interglacial periods, lasting about 10,000 years, our ancestors expanded their habitat and migrated to other hemispheres.
Until now evolutionary theories postulate that the dramatic leap in human evolution is the result of natural selection processes that occurred during the recent 2.5 million years. It was hypothesized that only the fittest and most intelligent among our ancestors would have survived these harsh conditions and would have been able to propagate. This hypothesis, however, cannot explain why the increase in brain size and other significant changes were limited to the ancestor of man and did not occur in other mammalian species. The concept presented in the following paragraphs can explain this phenomenon.
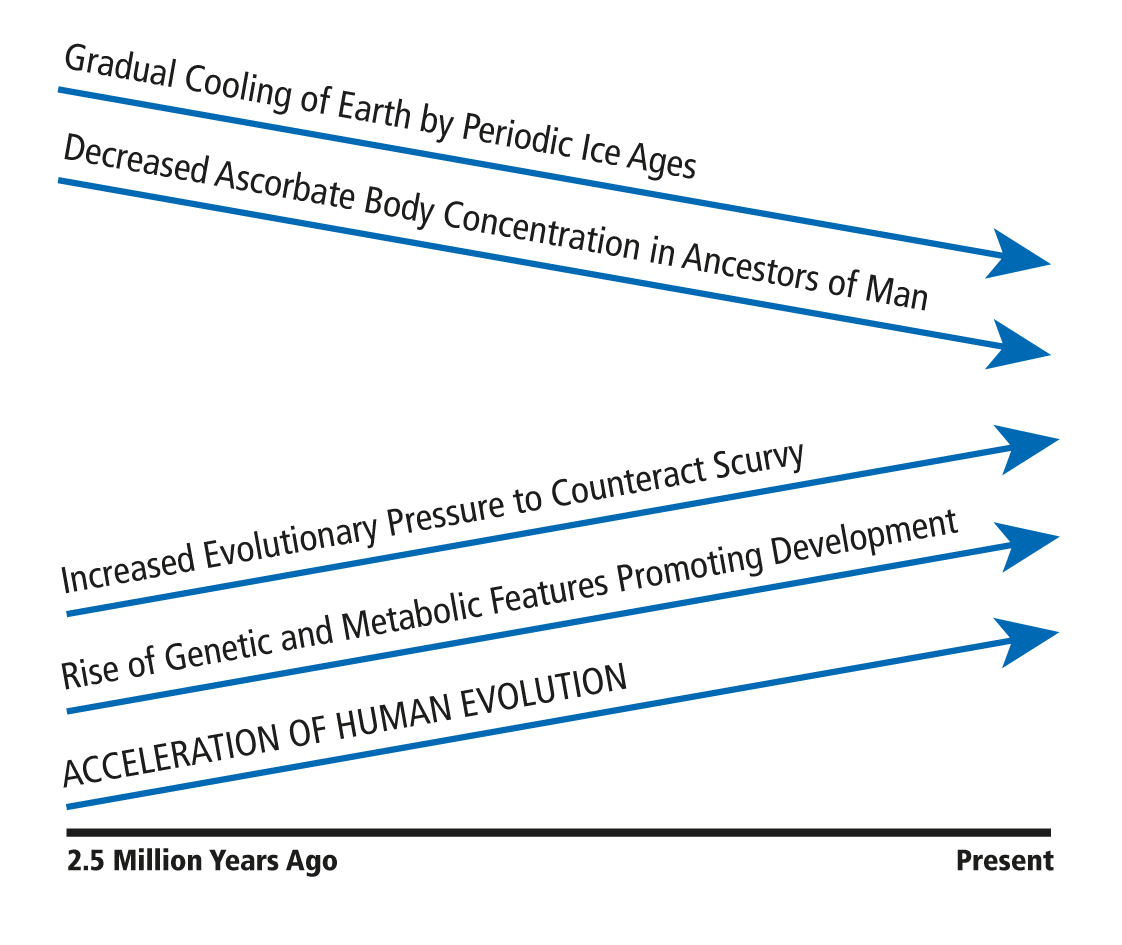
Figure 1.
The dramatic drop in temperature during the Ice Ages affected the vegetation on a global level. Scarce nutrition and frequent deficiency in vitamin and other essential nutrients affected all species equally. The ancestor of man shared with most other mammals genetic defects that rendered them susceptible to pellagra, pernicious anemia, beri-beri, and other diseases caused by nutritional deficiencies. Human metabolism, however, was set apart from the metabolism of other species by the inability for endogenous ascorbate production. While other species continued to manufacture ascorbate endogenously, generally at a rate of several grams per day compared to the human body weight, our ancestors’ body ascorbate concentration was limited by the low ascorbate intake in their diet. During the tens of thousands of years each glaciation period lasted ascorbate intake approximated zero and scurvy became the greatest threat to the evolutionary survival of man.
Scurvy – The Greatest Challenge For The Evolutionary Survival of Man
Scurvy is the result of total ascorbate depletion of the body and of a gross impairment of collagen and elastin synthesis. Scurvy is a fatal disease characterized by a virtual dissolution of the connective tissue throughout the body including the walls of the blood vessels. He sailors of earlier centuries died from scurvy, particularly from blood loss through the scorbutic vascular walls, within a few months. During the millennia of glaciation, billions of our ancestors died from scurvy particularly during the most recent Ice Ages, when they had migrated to climatically exposed hemispheres of the earth.
The death toll from scurvy was so enormous that our ancestors in many regions were virtually rendered to extinction. One example are the Neanderthals. These highly developed hominids living in many parts of Europe had became extinct during the last glaciation period. Which lasted from about 120,000 years ago to about 15,000 years ago. Neanderthal fossils reveal obvious signs of scurvy: frequent fractures of bones and disrupted growth of teeth.
Since scurvy was the greatest threat, the greatest pressure for the survival and the evolution of man was the need for genetic features able to counteract the fatal consequences of scurvy.
The Vascular Wall – the Focus of Genetic Adaptation
The focus of these countermeasures and of the associated genetic adaptation process was the vascular wall and the paramount need to counteract blood-loss through the scorbutic vascular wall. This adaptation process was characterized by a selective advantage of inherited features that rendered compensatory stability to the ascorbate-deficient vascular wall.¹
These genetic features comprise a multitude inherited metabolic disorders that can lead to the deposition of plasma constituents in the vascular wall, to proliferative responses of cellular systems in the vascular wall or by other mechanism resulting in a compensatory stabilization of the ascorbate-deficient vascular wall. By favoring these genetic features nature decided for the lesser of two evils: death from cardiovascular disease during adulthood rather than death from scurvy during infancy.
Ascorbate deficiency favored this genetic adaptation process against the fatal consequences of scurvy also in another way. Ascorbate is the strongest antioxidant in the body. Low ascorbate concentrations decrease the protection against oxidative damage of DAN and thereby increase the rate of genetic mutations.5 Ascorbate intake approximating zero during the millennia of glaciation initiated a form of ‘genetic roulette’ in our ancestors. The significantly increased genetic mutation rate greatly accelerated the genetic adaptation that favored not only countermeasures against scurvy but at the same time promoted human evolution. The more effective a genetic feature stabilizes the vascular wall during ascorbate deficiency the more important became this genetic feature as a metabolic promoter for the development of man.
Ascorbate Deficiency and Metabolic Promoters of Human Evolution
In general, all metabolic changes induced by ascorbate deficiency have, to a variable degree, affected organ development and differentiation during the evolution of man. Those metabolic factors that become available at increasing concentrations during ascorbate deficiency were metabolic promoters of evolution. Many of the metabolic changes induced by ascorbate deficiency are enhanced by a simultaneous deficiency in other essential nutrients such as niacin and riboflavin, which frequently interact synergistically with ascorbate. A deficiency of ascorbate, however, the strongest hydroxylating and reducing agent in the body, is the most important among them. In the following paragraphs I will focus on some metabolic interactions of ascorbate, which are important in the context of this publication.
An effective and therefore frequent mechanism counteracting scorbutic blood loss and therefore an important metabolic promoter of evolution was the elevation of plasma levels of lipid-rich ‘atherogenic’ lipoproteins. Low density lipoprotein (LDL), very low density lipoprotein (VLDL), and particularly lipoprotein(a) (Lp(a)) are found to be significantly elevated in humans compared to other species with endogenous ascorbate synthesis. Even more pronounced is the difference between man and ascorbate producing animals for the ratio between these ‘atherogenic’ lipoproteins and the ‘anti-atherogenic’ high-density lipoprotein (HDL). Atherogenic lipoproteins are characterized by a high content of lipids, e.g. cholesterol and fatty acids, and their elevated plasma concentration reflects an improved substrate supply for organ development and growth.
Ascorbate deficiency also leads to an increased availability of glucose. The increased availability of lipids and carbohydrates may in part be mediated by an increased corticotropin and cortisol release during ascorbate deficiency.6 Other mechanisms leading to an increased substrate supply in ascorbate-deficient conditions are reviewed elsewhere.1,6,7
However, improved metabolic substrate supply alone cannot explain the complex organ changes during evolution such as the development and differentiation of the human brain. These changes require an increased availability of metabolic factors involved in organ morphogenesis. Such metabolic factors are represented by a group of proteins called adhesive proteins. These proteins share a characteristic tripeptide sequence, arginine-glycine-aspartate (RGD), and they mediate the interaction between cellular systems and the extracellular matrix in a multitude of conditions such as organ differentiation, repair and growth (review in 8).
While ascorbate deficiency decreases the rate of synthesis for certain adhesive proteins such as collagen and fibronectin the production of certain other adhesive proteins such as fibrinogen and apo(a) is increased at low ascorbate concentrations. Of particular importance for organ development and differentiation during the evolution of man was apo(a).
Apoprotein(a) and Lipoprotein(a) – Decisive Metabolic Promoters of Human Evolution
Apo(a) and Lp(a) became major constituents in the metabolism of our ancestors after they had lost the ability to synthesize ascorbate. Lp(a), a unique combination of the adhesive protein apo(a) with an LDL particle, and apo(a) are quite likely the single most important metabolic promoters of evolution. One of the reasons for the selective evolutionary advantage of apo(a) and Lp(a) was their extraordinary effectiveness in counteracting scorbutic blood loss by their extracellular deposition in the ascorbate deficient vascular wall.9
The other reason for the selective advantage of the adhesive protein apo(a) is its contribution to the development of the human body during evolution. Beside the liver, where apo(a) is secreted as Lp(a), only two other organs were reported to have the ability for autonomous apo(a) production: the brain and the testes.¹º These two organs have determined critical evolutionary advantages: intelligence and fertility.
Apo(a) and Brain Development
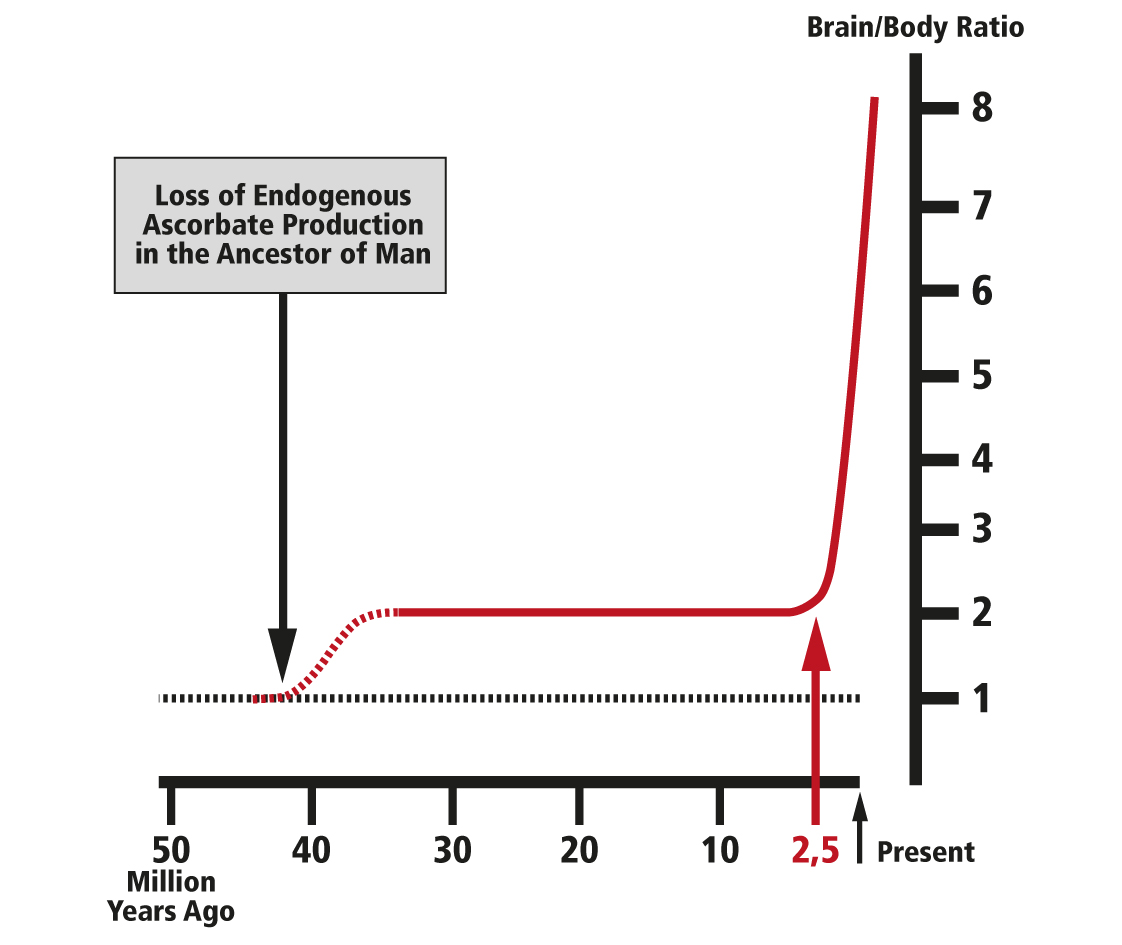
Figure 2.
A dramatic increase in brain size and differentiation during the evolution of man determined his dominant role today. Acquisition of language and ‘toolmaking’ have been proposed as factors responsible for the rapid encephalication. However, these factors are rather the result than the cause of increased brain size and intelligence. Moreover, these hypotheses leave open the decisive question of why a similar development did not occur in other mammals, which were exposed, to the same environmental conditions.
Apo(a) was an important metabolic factor in the development and the differentiation of the human brain during evolution. Like other adhesive proteins apo(a) contains an RGD tripeptide. RGD sequences are critically involved in the morphogenesis and differentiation of the central and peripheral nervous system (review in 8). During human evolution apo(a) synthesis rate in the brain continuously increased as the result of the genetic adaptation to counteract scurvy. Moreover, dietary deficiencies particularly of ascorbate and niacin during the millennia of glaciation let to a metabolic upregulation and a high apo(a) synthesis rate. It is therefore concluded that apo(a) has been a metabolic clue to the development of the human brain and the increase of intelligence during human evolution.
During this development apo(a) has been interacting with other adhesive proteins such as fibronectin and collagen. These adhesive proteins, however, cannot offer a clue to human evolution – they are present throughout the animal world and are not preferential features of human metabolism.
Apo(a) and Increased Fertility
Another decisive advantage during evolution was the improvement of fertility. This is even more remarkable since during evolution female reproductive physiology has lot an important signal, the estrus, and developed a concealed form of ovulation. Improved fertility was an important precondition for the development of concealed ovulation. Adhesive proteins are known to improve fertility by facilitating the interaction of sperm cells with egg cells as well as by mediating egg penetration (review in 8). Apo(a) was recently detected in human sperm ³ and increased apo(a) concentrations in the seminal fluid of our ancestors must have led to improved fertility. In conclusion, apo(a) was a metabolic clue in determining decisive advantages during the evolution of man: intelligence and fertility.
The role of apo(a) for the development of the body during human evolution was not confined to the brain and the testes. Elevated plasma levels of Lp(a) in newborns today¹¹ indicate an important role of apo(a) in development, differentiation and growth of the human body as a whole.
Plasma constituents reacting with antibodies against human apo(a) are also found in lower mammals, particularly in those with permanent or seasonal susceptibility to ascorbate deficiency such as the guinea pig and the hedgehog. These findings do not contradict the conclusions presented here. They rather underline human evolution as a multifactorial process with apo(a) and Lp(a) being of particular importance.
The Evolution of Man and Human Health Today
The mutation of a single gene encoding for a key enzyme in the conversion of glucose to ascorbate 40 million years ago in the ancestor of man became a two-sided sword. On one side this genetic mutation became the precondition for human evolution and was the decisive precondition why within the last 2.5 million years one species, man, became the dominant species on earth. On the other side this very same mutation left all descendants including over four billion people living today susceptible to scurvy and other characteristic diseases that are essentially unknown in animals with endogenous ascorbate production.
While scurvy is essentially unknown today, chronic insufficient dietary intake of ascorbate is widespread. Chronic ascorbate deficiency is the underlying cause for the most frequent diseases, diabetes, and other diseases. Millions of people die every year and millions more become disabled from these preventable diseases. Optimum dietary intake of ascorbate, particularly in combination with niacin, riboflavin, and other essential nutrients, should compensate for the genetic defect that lead to a cessation of endogenous ascorbate synthesis. The discoveries presented in this publication open the opportunity to greatly improve human health in this generation and future generations of mankind.
The Determining Principles of Human Evolution
The dramatic acceleration of human evolution during the recent 2.5 million years and the dominant role of humans on earth today are not the result of random selection during this period. Human evolution is the result of a unique combination of genetic, metabolic, environmental and dietary conditions.
The underlying genetic precondition for the evolution of man was a genetic mutation that occurred 40 million years ago in our ancestors: the loss of endogenous ascorbate production. This genetic mutation left all descendants dependent on dietary ascorbate intake and set their metabolism apart from other species which continued endogenous ascorbate production at an average daily rate of several thousand milligrams per day compared to the human bodyweight. The loss of endogenous ascorbate production resulted in a significant drop of body ascorbate concentrations in our ancestors. This fact may have triggered a first leap towards the evolution of man which occurred about 40 million years ago.
The Ice Ages, starting about 2.5 million years ago, became the environmental trigger condition for the evolution of man. Human evolution was particularly accelerated during the most recent Ice Ages, when our ancestors had migrated to the Northern Hemisphere and other parts of the world directly exposed to harsh climatic conditions. The cooling of the earth led to a decreased vegetation and to a limited availability of essential nutrients.
The dietary trigger condition for the evolution of man was an insufficient intake of vitamin C. During glaciation the ancestor of man shared with other mammals a limited food supply and a deficiency in most essential nutrients. Ascorbate deficiency, however, became a characteristic condition in the metabolism of our ancestors.
Scurvy was the greatest threat to the evolutionary survival of our ancestors particularly during the millennia of glaciation. While other mammals were protected during the Ice Ages from scurvy by their endogenous ascorbate synthesis, billions of our ancestors died from this disease.
The greatest evolutionary pressure during the evolution of man was the need for genetic and metabolic countermeasures to limit the fatal consequences of scurvy. These genetic countermeasures against scurvy had a selective evolutionary advantage over millions of generations.
In ascorbate deficiency the weakest sites of the body are the blood vessels, and hemorrhagic blood loss through the scorbutic vascular wall is a frequent cause of death in scurvy. The vascular wall became the focus of genetic countermeasures that protect the ascorbate-deficient walls against fatal blood loss.
Advantageous genetic features counteracting scurvy became the genetic and metabolic base for the evolution of man. The more effective a genetic or metabolic feature protected the vascular walls against scorbutic blood loss, the greater was its contribution to the development and differentiation of the human body during the evolution of man.
The single most important metabolic feature for the evolution of man was aporotein(a). In association with LDL, apo(a) became the most effective mechanism to stabilize the ascorbate-deficient vascular wall. As an adhesive protein expressed in the brain and the testes, apo(a) was involved in determining decisive evolutionary advantages: intelligence and fertility.
In general, all metabolic factors that become available at increased concentrations during ascorbate deficiency have to be considered a metabolic promoters of evolution. These metabolic factors include lipid substrates (cholesterol, triglycerides), and lipoproteins (Lp(a), LDL, VLDL).
The genetic adaptation was accelerated by the fact that ascorbate deficiency greatly favored the rate of genetic mutations. This increased mutation rate favored the selective evolutionary advantage of genetic and metabolic features counteracting scurvy an simultaneously promoting evolution.
The genetic mutation that resulted in the loss of endogenous ascorbate synthesis in the ancestor of man became a two-sided sword. On one side it let to a great death toll from scurvy and other diseases in the descendants; on the other side this mutation was a decisive genetic precondition for man to become the dominant species on earth.
The critical role of ascorbate deficiency during human evolution has immediate implications for the health of human beings today. While scurvy is essentially unknown today, chronically insufficient dietary intake is widespread. Ascorbate deficiency is a precondition for the most frequent human diseases today, including cardiovascular diseases, diabetes, and many other diseases. These diseases are essentially unknown in animals producing high amounts of ascorbate endogenously and they can be prevented in humans by optimum dietary ascorbate supplementation.
Conclusion
In this paper I presented decisive missing pieces in the puzzle of human evolution. The remaining questions may largely be answered by a new scientific discipline: metabolic anthropology. The solution to the puzzle of human evolution and its direct implications for human health today is a modest contribution by an individual scientists, but it may turn out to be a major step for mankind.
References
Rath, M., Pauling, L. A unified theory of human cardiovascular disease leading the way to the abolition of this disease as a cause for human mortality. Journal of Orthomolecular Medicine 1992; 7: 5-15
Rath M, Pauling L, Hypothesis: Lipoprotein (a) is a surrogate for ascorbate. Proceedings of the National Academy of Sciences USA 1990; 87: 6204-6207
Rath M, Pauling L. Apoprotein(a) is an adhesive protein. Journal of Orthomolecular Medicine 1991; 6:139-143.
Calvin W. The ascent of mind, Ice Age climates and the evolution of intelligence. Bantam Books, New York, Toronto London, Sydney, Auckland 1991.
Fraga CG, Motchnik PS, Shigenaga MK, Helbock HJ. Jacob RA, Ames BN. Ascorbic acid protects against endogenous oxidative DANN damage in human sperm. Proceedings of the National Academy of Sciences USA 1991; 88: 11003-11006.
Clementson CAB: Vitamin C, Volume I-III. Florida: CRC Press, Inc. 1989.
Burns JJ, Rivers JM, Machlin LJ, eds. Third Conference on Vitamin C, Annals of the New York Academy of Sciences 1987, 498.
Hynes R.O.: Fibronectins. Springer-Verlag Series in Molecular Biology, Springer-Verlag, New York 1990.
Rath, M., Pauling, L. Solution to the puzzle of human cardiovascular disease: its primary cause is ascorbate deficiency leading to the deposition of lipoprotein (a) and fibrinogen/fibrin in the vascular wall. Journal of Orthomolecular Medicine 1991; 6:125-134.
Tomlinson J.E., McLean J.W., Lawn R.M.: Rhesus monkey apolipoprotein(a). Sequence, evolution and sites of synthesis. Journal of Biological Chemistry 1989; 26664: 5957-5965.
Van Biervliet JP, Labeur C, MNichiels G, Usher DC, Rosseneu M. Lipoprotein(a) profiles and evolution in newborns. Atherosclerosis 1991, 86: 173-181.
Eccles JC. The human mystery. Routledge & Kegan Paul, London, Boston, Melbourne and Henley 1984.

{kind=link}